Pharmacophore an International Research Journal
AN IN- SILICO STUDIES FOR IMMUNOMODULATORY POTENTIAL OF PHYTOCONSTITUENTS FROM A NATURALLY OCCURRING HERB NIGELLA SATIVA
Snehal Kashid1, Ashish Suttee1*, Prasad Kadam2, Gopal Lal Khatik3, Ramesh Kasarla4
|
|
|
ABSTRACT
Nigella sativa L. belongs to the family of Ranunculaceae, commonly known as Black Cumin or Kalonji. It is an annual herb used as a food ingredient and spice. N. Sativa is approved by the US Food and Drug Administration (FDA) as food and by the Flavor and Extract Manufacturers Association (FEMA) as generally recognized as safe (GRAS). We aimed to identify the potent immunomodulatory phytoconstituents of N. Sativa using the in-silico studies like molecular docking. For docking assessment, two proteins 1M48, and 1P9M were used for ten active constituents (Carvacrol, p-Cymene, Dithymoquinone, Gamma himachalene, Limonene, Nigellamine C, Nigellicine, Nigellidine, alpha-Pinene, and Thymoquinone) are selected. In-silico ADME studies were also performed. The molecular docking study is done using AutodockVina 1.1.2 and visualized by BIOVIA Discovery Studio. Curcumin is chosen as the standard of natural origin, while dexamethasone and salazosulfapyridine are synthetic standards. All the phytoconstituents showed good binding affinities with 1M48 (IL-2) protein in the range of -4.3 to -7.3 kcal/mol, while on 1P9M (IL-6) protein, the binding affinities observed to be in the range of -5.2 to -9.8 kcal/mol. The in-silico ADME studies showed that these phytoconstituents have better druggability and no cytotoxicity. The current study showed that the immunomodulatory properties of N. Sativa are from Nigellamine C.
Keywords: In silico, Immunomodulatory, Protein-ligand, Nigella sativa, Phytoconstituents, Interleukins
Introduction
Defending the host from the pathogenic destructive microorganism in vertebrates is the immune system. This protective mechanism of the body is extremely necessary for the investigation of host tissue integrity. The chief distinctive feature of immunomodulatory agents is to augment or diminution the immune response generated by host immune cells [1, 2]. Natural ingredients can control and cure acute, chronic disease infectious diseases, etc. the major advantage of using natural constituents is they have a novel structure, multi-target, safe and effective remedy [3, 4]. A notable rise in the occurrence and austerity of adverse drug reactions observed with several synthetic drugs ultimately boosted substantial interest in natural remedies [5]. From antique times, various traditional systems of medicine naming Ayurveda, Siddha, Tibb, Unani, and the Chinese system (TCM) substantiate several natural products, plants, and extracts possessing the ability to modulate the immune system [6, 7]. Considerable innovation and research were carried out on establishing and developing novel immunomodulatory agents. Amongst the few top-ranked evidence-based herbal medicines, Nigella sativa (Ranunculaceae) has a special place [8, 9]. It is native to Middle Europe, and Western Asia. N. Sativa is immersed in a rich historical and religious context. N.Sativa is an annual blossoming herb with erect stems,20-30 cm in height and green to dark green [10, 11]. The flowers are delicate white and turns blue at the time of maturity with 5 to 10 petals [11]. The fruits are inflated capsulesenclosed black colored, oval seeds with an approximate size of 1-3 mm [12]. Nigella sativa consists of fixed oil, volatile oil, alkaloids, proteins, saponin, flavonoids, phenolic acids, and tannins [6, 13, 14]. There are different active phytoconstituents present in N. Sativa, such as thymoquinone, thymohydroquinone, dithymoquinone, cymene, carvacrol, pinene, terpineol, limonene, anethol, hederin, citronellol, nigellicine, nigellidine, nigellamines, etc. [6, 15-18]. Seeds of Nigella sativa have been increasingly reported as a successful therapeutic agent. It showed antimicrobial, antibacterial, antifungal, antiparasitic, anticancer, anti-inflammatory, and immunomodulatory activities, helpful in atherosclerosis, asthma, cough, bronchitis, influenza, and eczema. Traditionally used as a diuretic, for headache, fever, bronchial disorders, insomnia, and titanus [19-24].
In recent years molecular docking becoming the key tool in computer-aided drug design for identifying novel ligands, binding affinity, and behavior of small molecules at the binding site of the target protein. With the help of molecular docking, we can be predicting the orientation of ligand bound to protein, protein-ligand, or protein-protein binding affinities, via various interactions [25-27]. The sum of these interactions between protein-ligand or protein-protein approximately represents the binding potential in the form of a docking score [26, 27]. The present study aimed to explore in silico interaction of them phytoconstituents with protein 1M48 (IL-2) and 1P9M (IL-6). 1M48 possesses a total structure weight of about 31.77 kDa while 1P9M has a total structure weight of 78.25 kDa.
Materials and Methods
Molecular Docking
Molecular docking is an important computational tool to identify the binding interaction at the target site and develop the pharmacophore [28].
Software and Database
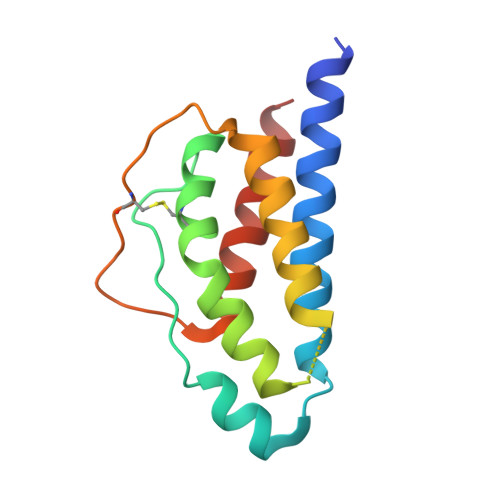
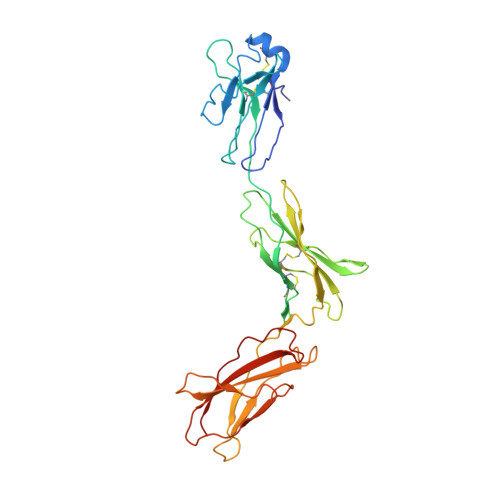
Autodock_vina version 4.2.6 (https://vina.scripps.edu) software approaches were used to predict the in-silico interaction between active compounds of N. Sativa and immunomodulatory proteins like 1M48 (Human interleukin-2 or IL-2 protein) and 1P9M (Human interleukin-6 or IL-6 protein) observed in Figure 1 [29]. BIOVIA Discovery Studio was downloaded from https://discover.3ds.com/ for visualization [30]. Open bable 3.1.1 was downloaded from http://openbabel.organd used to convert SDF to PDB format [31]. The calculations were performed on Windows 10 operating system.
Ligand Selection
Ten phytoconstituents i.e. Carvacrol, p-Cymene, Dithymoquinone, Gamma himachalene, Limonene, Nigellamine C, Nigellicine, Nigellidine, alpha-Pinene, and Thymoquinone are selected as the ligand (Figure 2). The 2D and 3D structures of these ligands in SDF format were downloaded from https://pubchem.ncbi.nlm.nih.govto dock against Interleukins. Natural phytoconstituent curcumin while synthetic compounds dexamethasone and salazosulfapyridine were selected as standard. Ligand preparation was performed in open bable 3.1.1 (http://openbabel.org) [31].
Preparation of Protein
The three-dimensional (3D) structure of the protein was extracted from the protein data bank (PDB) (http://www.rcsb.org). The extracted protein was PDB ID: 1M48, PDB ID: 1P9M, and its chain were shown in Figures 1a and 1b, respectively. BIOVIA Discovery Studio was utilized for protein preparation [30]. All water molecules & het-atoms which responsible for increased resolution were deleted. Observe active sites, protein groups, and ligand groups.
Target and Ligand Optimization
In molecular docking, an optimization algorithm was utilized to find a suitable binding pose of a ligand against a protein target to show high binding affinity. This algorithm plays a dynamic role in determining the docking accuracy. A suitable binding pose was determined with autodock vina (version 4.2.6) and visualized with the help of BIOVIA Discovery Studio [29, 30].
Analysis of Target Active Binding Sites
The active site of protein comprised of amino acids was determined by the CASTp tool (Computed Atlas for Surface Topography of Proteins) (Table 1) (http://sts.bioe.uic.edu) [32]. Pharmacophore development of targeted protein PDB ID: 1M48 and PDB ID: 1P9M was analyzed with the help of BIOVIA Discovery Studio. After protein preparation, a single ligand is selected, and the receptor-ligand interaction is observed; the binding site and the expansion or SBD site sphere are edited. For PDB 1M48, the size of the sphere was 20 Å. On the contrary, for 1P9M, it was 80 Å. Afterward processing of protein, the ligand group was deleted, Polar hydrogen atoms and the Kollman charges were added, and the file was saved in PDB format [33, 34].
Molecular Docking Analysis
A docking study was employed to study the interaction between protein and ligand. The selected protein for docking plays roles with several intermediates, such as PDB ID: 1M48 (IL 2), and PDB ID: 1P9M (IL-6). Proteins were converted from PDB to PDBQT format with the help of the AutodockVina tool. The configuration files were prepared for both proteins Further, using a command prompt, docking was processed with a ligand file. The exhaustiveness of the global search was set to eight, and the maximum number of binding modes was nine. For the analysis of our ligand file, we used Biovia Discovery Studio to define ligand interaction [29, 30].
ADME Profiling
From the retrieved data of molecular docking of ten phytocompounds from Nigella sativa against target proteins, active constituents Carvacrol, p-Cymene, Dithymoquinone, Nigellicine, Nigellidine, alpha-Pinene, and Thymoquinone were considered for ADME properties using the Swiss ADME (http://www.swissadme.ch/index.php) to predict pharmacokinetic properties of phytoconstituents [35, 36].
Results and Discussion
Several protein structures were available for studying the immunomodulatory potential of the phytoconstituents from N. Sativa. We have selected the 1M48 (Human interleukin-2 or IL-2 protein) and 1P9M (Human interleukin-6 or IL-6 protein) and downloaded them from the RCSB protein data bank for the molecular docking study (Figure 1). Molecular docking efficiently predicts binding affinities, amino acid involvements, and protein-ligand complexes and became one of the important strategies during drug discovery [37-41].
|
|
|
|
a) |
b) |
|
Figure 1. 3D structure of 1M48 (a) and 1P9M (b) proteins |
|
The protein code 1M48 is a crystal structure of human IL-2 complexed with (R)-N-[2-[1-(aminoiminomethyl)-3-piperidinyl]-1-oxoethyl]-4-(phenylethynyl)-L-phenylalanine methyl ester and 1P9M is a crystal structure of the hexameric human IL-6/IL-6 alpha-receptor/gp130 complex. Ten known phytoconstituents i.e., Carvacrol, p-Cymene, Dithymoquinone, Gamma himachalene, Limonene, Nigellamine C, Nigellicine, Nigellidine, alpha-Pinene, and Thymoquinone have used a ligand (Figure 2). To compare them, natural standards like curcumin and synthesis standards like dexamethasone and salazosulfapyridine were used (Figure 3). A flow of the process is selection, retrieving of proteins and ligands, molecular docking, and ADME study to find potential immunomodulatory phytoconstituent.
|
|
|
Figure 2. Selected phytoconstituents from N. sativa. |
|
|
|
a) Herbal |
|
|
|
b) Synthetic |
|
Figure 3. Structures of the standard used for comparison. |
Table 1. Binding pockets of protein along with sequence
|
Binding pocket |
Sequence |
|
1M48 |
|
|
1P9M |
|
Table 1 Indicates the binding pockets in red color and highlighted residue sequence forming the binding pocket obtained from the CASTp tool [32].
The molecular docking results, including binding affinities and interactions of ten phytoconstituents and three standards, are presented in Tables 2 and 3 to the targeted protein 1M48 and 1P9M respectively, with 2D and 3D presentation.
The ten phytoconstituents interacted individually with chain A 1M48 protein (presenting human IL-2) with LEU72, ARG38, ARG81, PRO82, ARG38, LYS43, PHE42, ALA73, LYS35, LYS35, MET39, and PRO65 amino acid residues. Among all the screened phytoconstituents, Nigellamine C showed a better binding affinity of -6.3 kcal/mol, which is close to the standard curcumin (-7.0 kcal/mol), and salazosulfapyridine (-7.3 kcal/mol). While alpha-pinene showed the lowest binding affinity of -4.3 kcal/mol.
Table 2. Interactions of different phytoconstituents with protein 1M48





The study showed that these phytoconstituents interacted with the Chain B of 1P9M protein (presenting human IL-6) and bound with LEU57, ALA56, ALA58, LYS54, ARG168, ARG30, LEU33 amino acid residues. In addition to Chain B, these also interact with Chain C of 1P9M protein and are bound with PHE168, PHE134, GLN190, ASN136, ARG233, GLU278, and PHE279 amino acid residues. Among the all screened phytoconstituents, Nigellamine C shows the highest binding affinity of -9.8 kcal/mol, which is far better than standards like curcumin (-6.7 kcal/mol), dexamethasone (-8.1 kcal/mol) and Salazosulfapyridine (-7.7 kcal/mol). At the same time, Thymoquinone showed the lowest binding affinity of -5.1 kcal/mol (Table 3).
Table 3. Several ligand interactions against protein 1P9M




A bar diagram (Figure 4) is drawn for a better comparison of the binding affinities of the different phytoconstituents and standards on both the IL-2 protein (1M48) and IL-6 protein (1P9M). This resulted in the most promising anti-inflammatory and immunomodulatorphytoconstituent, Nigellamine C, which could inhibit IL-2 and IL-6. Such correlation can be made from previous literature reports, suggesting the role of IL-2 and IL-6. The interleukin-2 (IL-2) formed by activated T cells through immune response helps transform naïve T cells into effector T cells [42]. It is synthesized as 153 amino acid polypeptide as a precursor and then matured IL2 which is composed of 133 amino acids with 15.5 kDa [43]. IL-2 promotes further growth and development of native immune cells and promotes survival and cell proliferation [42, 44, 45]. IL 2 is used in several inflammatory and autoimmune diseases recently, it was approved by US Food and Drug Administration (FDA) in treating melanoma and kidney cancer as immunotherapy [42, 46, 47].
Docking interactions of these phytoconstituents along with protein are presented below. Tables 2 and 3 indicate the active site of protein 1M48 and 1P9M respectively, where a total of 10 bioactive constituents interact individually. Chain A residue of 1M48 bound with LEU72, ARG38, ARG81, PRO82, ARG38, LYS43, PHE42, ALA73, LYS35, LYS35, MET39, and PRO65. Among all the tested phytoconstituents salazosulfapyridine shows the highest (-7.3) docking score while pinene shows the lowest (-4.3) docking score presented in Table 2. The B chain of 1P9M bound with LEU57, ALA56, ALA58, LYS54, ARG168, ARG30, LEU33 additionally chain C bound with PHE168, PHE134, GLN190, ASN136, ARG233, GLU278, and PHE279, between all phytoconstituents nigellamine C shows the highest binding affinity (-9.8) on the other hand thymoquinone shows lowest binding affinity (-5.1) represented in Table 3.
|
|
|
Figure 4. Protein binding affinity (kcal/mol) with active constituents of Nigella sativa |
Majorly Interleukin-6 is secreted by leukocytes, monocytes, and macrophages. Human Interleukin 6 (IL-6) consists of 212 amino acids, 28-amino-acid signal peptides, and 21–26 kDa [48]. IL 6 is a cytokine with pleiotropic functions produced in response to tissue damage and regeneration, hematopoietic regulation, sustaining tumorigenesis, and infections [49]. IL6 acts as pro-inflammatory and anti-inflammatory as cytokine and myokine, respectively. Adding osteoblasts secrete IL-6 to stimulate bone formation while Smooth muscle cells from the middle layer of the artery secret IL6 as an anti-inflammatory myokine. Overproduction of IL6 causes the risk of inflammatory bowel disease, rheumatoid arthritis, and juvenile idiopathic arthritis [40, 41, 50].
Further after molecular docking studies, we performed the in-silico ADME studies of Carvacrol, p-Cymene, Dithymoquinone, Nigellicine, Nigellidine, alpha-Pinene, and Thymoquinone using the Swiss ADME (http://www.swissadme.ch/index.php) [35, 36]. The results showed that all these phytoconstituents showed no cytotoxicity and metabolic interferences with membrane receptors or Cyps. This suggests that these could be better drug candidates with good pharmacokinetic profiles. Most of these have good water solubility. Few have low GI absorption and BBB penetration. Also mostly followed Lipinski’s rule of druggability.
In the present study, the phytoconstituents showed the interaction only with Chain A of 1M48 (IL-2) protein while involving Chain B and C of 1P9M (IL-6) protein. IL-2 interacts via LEU72, PHE42, and ARG38 through a hydrophobic bond, although IL-6 is bound via ARG168, GLN190, and ASN136, forming a hydrogen bond. The most promising Nigellamine C showed binding affinities of -6.3 kcal/mol and -9.8 kcal/mol with IL-2 and IL-6, respectively.
Conclusion
The present study examines the immunomodulatory potential of ten phytoconstituent of Nigella sativa on two immunomodulatory target proteins through in-silico studies, including molecular docking by AutodockVina and ADME by Swiss AMDE. These phytoconstituents were Carvacrol, p-Cymene, Dithymoquinone, Gamma himachalene, Limonene, Nigellamine C, Nigellicine, Nigellidine, alpha-Pinene, and Thymoquinone. The molecular docking and ADME studies suggested that Nigellamine C would be the most promising immunomodulator. However, further studies on these phytoconstituents must establish a novel, safe and effective immunomodulators.
Acknowledgments: The research facilities from the School of Pharmaceutical Sciences, Lovely professional university Panjab; and SPMs College of Pharmacy, Akluj, Maharashtra are highly acknowledged.
Conflict of interest: None
Financial support: None
Ethics statement: Authers were personally involved in presented research work and take whole responsibility for its content.
1. Pasala PK, Reddy LS, Silvia N, Reddy YD, Sampath A, Dorababu N, et al. Molecular docking and in vivo immunomodulatory activity of Albizia procera bark on doxorubicin induced immunosuppressive rats. J King Saud Univ Sci. 2022;34(3):101828.
2. Delves PJ, Martin SJ, Burton DR, Roitt IM. Roitt's essential immunology. John Wiley & Sons; 2017.
3. Maurya SK, Mishra R. Molecular docking studies of natural immunomodulators indicate their interactions with the CD40L of CD40/CD40L pathway and CSF1R kinase domain of microglia. J Mol Model. 2022;28(4):1-3.
4. Jin X, Liu MY, Zhang DF, Zhong X, Du K, Qian P, et al. Natural products as a potential modulator of microglial polarization in neurodegenerative diseases. Pharmacol Res. 2019;145:104253.
5. Hwang J, Yadav D, Lee PC, Jin JO. Immunomodulatory effects of polysaccharides from marine algae for treating cancer, infectious disease, and inflammation. Phytother Res. 2022;36(2):761-77.
6. Khan S, Ali M, Albratty MM, Najmi AY, Azeem U, Khan SA, et al. Nigella sativa: From chemistry to medicine. Pharmacol Ther Appl. 2022;(2):29-62.
7. Imran M, Khan SA, Alshammari MK, Alkhaldi SM, Alshammari FN, Kamal M, et al. Nigella sativa L. and COVID-19: A Glance at The Anti-COVID-19 Chemical Constituents, Clinical Trials, Inventions, and Patent Literature. Molecules. 2022;27(9):2750.
8. Zaidi Z. A Brief Study of Nigella sativa (Kalonji): A Unique Immunomodulator of Unani Medicine. Issues Dev Med Med Res. 2022;4:102-9.
9. Jakhmola Mani R, Sehgal N, Dogra N, Saxena S, Pande Katare D. Deciphering underlying mechanism of Sars-CoV-2 infection in humans and revealing the therapeutic potential of bioactive constituents from Nigella sativa to combat COVID19: in-silico study. J Biomol Struct Dyn. 2022;40(6):2417-29.
10. Dalli M, Bekkouch O, Azizi SE, Azghar A, Gseyra N, Kim B. Nigella sativa L. Phytochemistry and Pharmacological Activities: A Review (2019–2021). Biomolecules. 2022;12(20):1-37.
11. Forouzanfar F, Bazzaz BS, Hosseinzadeh H. Black cumin (Nigella sativa) and it's constituent (Thymoquinone): a review on antimicrobial effects. Iran J Basic Med Sci. 2014;17(12):929-38.
12. Hebidi M. Contribution à l’étude de la graine de nigelleou cumin noir Nigella sativa L. Master’s Thesis, Aix-Marseille Université, Marseille, France, 2019.
13. Dalli M, Azizi SE, Benouda H, Azghar A, Tahri M, Bouammali B, et al. Molecular Composition and Antibacterial Effect of Five Essential Oils Extracted from Nigella sativa L. Seeds against Multidrug-Resistant Bacteria: A Comparative Study. Evid Based Complement Altern Med. 2021;9(1).
14. Ahmad MF, Ahmad FA, Ashraf SA, Saad HH, Wahab S, Khan MI, et al. An updated knowledge of Black seed (Nigella sativa Linn.): Review of phytochemical constituents and pharmacological properties. J Herb Med. 2021;25:100404. doi:10.1016/j.hermed.2020.100404
15. Salehi B, Quispe C, Imran M, Ul-Haq I, Živković J, Abu-Reidah IM, et al. Nigella plants–Traditional uses, bioactive phytoconstituents, preclinical and clinical studies. Front Pharmacol. 2021;12:625386.
16. Khan LM, Karim S. Pharmacological basis of Thymoquinone as a putative adjuvant anticonvulsant-a systematic review. Int J Pharm Res Allied Sci. 2020;9(3):131-42.
17. Alshwyeh HA, Aldosary SK, Ilowefah MA, Shahzad R, Shehzad A, Bilal S, et al. Biological Potentials and Phytochemical Constituents of Raw and Roasted Nigella arvensis and Nigella sativa. Molecules. 2022;27(2):550.
18. Siddiqui S, Upadhyay S, Ahmad R, Gupta A, Srivastava A, Trivedi A, et al. Virtual screening of phytoconstituents from miracle herb nigella sativa targeting nucleocapsid protein and papain-like protease of SARS-CoV-2 for COVID-19 treatment. J Biomol Struct Dyn. 2022;40(9):3928-48.
19. Shahid MA, Rahim A, Chowdhury MA, Kashem MA. Development of antibacterial nanofibrous wound dressing and conceptual reaction mechanism to deactivate the viral protein by Nigella sativa extract. Adv Tradit Med. 2022;22(2):283-91.
20. Kabir Y, Akasaka-Hashimoto Y, Kubota K, Komai M. Volatile compounds of black cumin (Nigella sativa L.) seeds cultivated in Bangladesh and India. Heliyon. 2020;6(10):e05343.
21. Islam MT, Guha B, Hosen S, Riaz TA, Shahadat S, Da Rocha Sousa L, et al. Nigellalogy: a review on Nigella sativa. MOJ Bioequiv Availab. 2017;3(6):00056.
22. Forouzanfar F, Bazzaz BS, Hosseinzadeh H. Black cumin (Nigella sativa) and its constituent (thymoquinone): a review on antimicrobial effects. Iran J Basic Med Sci. 2014;17(12):929-38.
23. Shafodino FS, Lusilao JM, Mwapagha LM. Phytochemical characterization and antimicrobial activity of Nigella sativa seeds. PloS one. 2022;17(8):e0272457.
24. Salehi B, Quispe C, Imran M, Ul-Haq I, Živković J, Abu-Reidah IM, et al. Nigella plants–Traditional uses, bioactive phytoconstituents, preclinical and clinical studies. Front Pharmacol. 2021;12:625386.
25. Pagadala NS, Syed K, Tuszynski J. Software for molecular docking: a review. Biophys Rev. 2017;9(2):91-102.
26. Taguchi YH, Turki T. A new advanced in silico drug discovery method for novel coronavirus (SARS-CoV-2) with tensor decomposition-based unsupervised feature extraction. PloS one. 2020;15(9):e0238907.
27. Prabhu DS, Rajeswari VD. In vitro and in silico analyses of Vicia faba L. on Peroxisome proliferator–activated receptor gamma. J Cell Biochem. 2018;119(9):7729-37.
28. Dhameliya TM, Nagar PR, Gajjar ND. Systematic virtual screening in search of SARS CoV-2 inhibitors against spike glycoprotein: pharmacophore screening, molecular docking, ADMET analysis and MD simulations. Mol Divers. 2022:1-8. doi:10.1007/s11030-022-10394-9
29. Trott O, Olson AJ. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem. 2010;31(2):455-61.
30. BIOVIA, Dassault Systèmes. Comprehensive modeling and simulations for life sciences. Biovia Discovery Studio. 2016;1.
31. O'Boyle NM, Banck M, James CA, Morley C, Vandermeersch T, Hutchison GR. Open Babel: An open chemical toolbox. J Cheminform. 2011;3(1):1-4.
32. Tian W, Chen C, Lei X, Zhao J, Liang J. CASTp 3.0: computed atlas of surface topography of proteins. Nucleic Acids Res. 2018;46(W1):W363-W7. doi:10.1093/nar/gky473
33. Xue Q, Liu X, Russell P, Li J, Pan W, Fu J, et al. Evaluation of the binding performance of flavonoids to estrogen receptor alpha by Autodock, Autodock Vina and Surflex-Dock. Ecotoxicol Environ Saf. 2022;233:113323.
34. Xia B, Luo M, Pang L, Liu X, Yi Y. Lipopeptides against COVID-19 RNA-dependent RNA polymerase using molecular docking. Biomed J. 2021;44(6):S15-24.
35. Daina A, Michielin O, Zoete V. SwissADME: a free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci Rep. 2017;7(1):1-3.
36. Daina A, Zoete V. A boiled‐egg to predict gastrointestinal absorption and brain penetration of small molecules. Chem Med Chem. 2016;11(11):1117-21.
37. Chikhale RV, Sinha SK, Khanal P, Gurav NS, Ayyanar M, Prasad SK, et al. Computational and network pharmacology studies of Phyllanthus emblica to tackle SARS-CoV-2. Phytomed Plus. 2021;1(3):100095.
38. Tan L, Xu Q, Shi R, Zhang G. Bioinformatics analysis reveals the landscape of immune cell infiltration and immune-related pathways participating in the progression of carotid atherosclerotic plaques. Artif Cells Nanomed Biotechnol. 2021;49(1):96-107. doi:10.1080/21691401.2021.1873798
39. Aloufi BH. Screening of Phytochemicals against Osteoporosis: Molecular Docking and Simulation-Based Computational Approaches. Int J Pharm Res Allied Sci. 2022;11(1):87-9.
40. Ganeshpurkar A, Saluja A. In silico interaction of hesperidin with some immunomodulatory targets: A docking analysis. Indian J Biochem Biophys. 2019;56(1):28-33.
41. Ganeshpurkar A, Saluja A. In silico interaction of rutin with some immunomodulatory targets: a docking analysis. Indian J Biochem Biophys. 2018;(55):88-94.
42. Tonya CW, Elvira LL, Kenneth JOB, William CSC, Steven MD. Microenvironment and Lung Cancer. IASLC Thorac Oncol. 2018;121:8.
43. Asao H. Interleukin-2. Encycl Endocr Dis. 2004:60-3.
44. Kattner AA. A finger in every pie e The versatility of chemokines. Biomed J. 2022;45(3):427-31.
45. Jiang T, Zhou C, Ren S. Role of IL-2 in cancer immunotherapy. Oncoimmunology. 2016;5(6):e1163462.
46. Mizui M. Natural and modified IL-2 for the treatment of cancer and autoimmune diseases. Clin Immunol. 2019;206:63-70.
47. Gougeon ML, Chiodi F. Impact of γ‐chain cytokines on T cell homeostasis in HIV‐1 infection: therapeutic implications. J Intern Med. 2010;267(5):502-14.
48. Velazquez-Salinas L, Verdugo-Rodriguez A, Rodriguez LL, Borca MV. The role of interleukin 6 during viral infections. Front Microbiol. 2019;10:1057.
49. Enayatjazi M, Esfarjani F, Reisi J, Moshtaghian SJ. Studying the effect of garlic consumption and endurance training on serum levels of some pro-and anti-inflammatory cytokines in female mice with breast cancer-A randomized trial. Int J Prev Med. 2022;13:38.
50. Yoshida Y, Tanaka T. Interleukin 6 and rheumatoid arthritis. Biomed Res Int. 2014; 2014.