Pharmacophore an International Research Journal
ANTIAGING EFFECT OF TEA EPIGALLOCATECHIN GALLATE AND ITS ROLE IN MODIFYING AGING EPIGENETICS: A SYSTEMATIC REVIEW
Reema A. Alyamani1*, Nada A. Almatrafi1
|
|
|
ABSTRACT
Increased life expectancy turns the public into aged societies, and aged individuals are expected to reach 2 billion of the world’s population. Aging is a time accumulating decline in the body's biological function. An important aging hallmark is epigenetic alteration. Tea is a rich source of Epigallocatechin Gallate which has a role in aging epigenetics modulation. This review is conducted to clarify and derive a more precise description of the association between Epigallocatechin Gallate (EGCG) and epigenetic alteration as an aging hallmark. The search was conducted in four databases (PubMed, Science Direct, Scopus, and Google Scholar) for peer-reviewed articles published between January 2011 and December 2020. Studies were selected according to the association between EGCG and regulation of aging-related epigenetic marks in mice animal models following the PRISMA statement checklist. The role of EGCG in muscle aging is the modification of miRNA, which decreases gene expression and proteins responsible for muscle wasting, another role of EGCG in aging through increasing AcH3K9 and HDAC1 lower expression, which improves cardiac diastolic function. EGCG is one of the epigenetics modifying phytochemicals in aging; according to the aging theory, histone acetylation restoration by HDAC 1 inhibitors, and there is a (gero-miRNAs) regulating aging-associated genes activity.
Keywords: Medicinal plant, Oxidative stress, Free radicals, Testicular toxicity, Aging, Polyphenols
Introduction
Aging
To our knowledge, this review is the first on the EGCG effect on epigenetic alteration as a hallmark of aging [1]. Increased life expectancy in both developed and developing countries, turning their public into aging societies, the United Nations defines the population over the age of 65 are about 7% of the whole population [2, 3]. In 2050 it is expected that people aged 60 years and older will reach 2 billion of the world's population [4]
Statistics highlight the increasing number of aged populations, therefore, needs more consideration for aging [5, 6]. Although aging is a time accumulating decline in biological function, several cellular process alterations, and DNA gene transcription and translation modulation changes could be due to factors some of them are genomic instability, telomere attrition, cellular senescence [7-9], and epigenetics alteration [10-12].
Characteristics of aging are inflammations, sarcopenia, and multiple disease susceptibility, including cardiovascular diseases, neurodegenerative diseases, cancers, chronic diseases, and other metabolic diseases; these features could be induced by bad lifestyle habits, poor diet quality, and physical inactivity [3, 13].
People age differently; the characteristics of aging are not for all aged individuals [14, 15]. Notably, centenarians people aged 100 years or older seem to have slower aging than others, indicating a difference between people in the aging process [16].
The research investigates the aging process at the body’s cellular and nuclear levels to find a solution that could reverse the side effects of the aging process since it has serious side effects such as multiple diseases and frailty [17, 18]. One of the important aging hallmarks is epigenetic alteration [11, 19]. As people age differently, the concept of epigenetic aging acceleration (EAA) has been invented to describe the difference between predicted DNA epigenetic age and chronological age [16, 20]. The EAA manifests through aging disease onset diagnosis in older adults as an aging consequence due to internal and external factors [21-23] build in the body and affect health for a long time [16].
Epigenetics
DNA in the cells contains cell identity, function, and instruction [24]. Epigenetics regulates the interactions between the genome and the external environment altering gene expression in response to environmental cues [25]. Epigenetics controls DNA accessibility and affects gene transcription without changing DNA sequence [25, 26].
The genetic material in cells is wrapped around histones protein; this DNA-protein complex is coiled into the shape of chromatin, and they are all condensed in the nucleus; DNA chromatin structure is highly organized to regulate genetic function [24, 27]. Epigenetics regulation includes histone modifications, DNA methylation [28], and non-coding RNA (ncRNA) [25, 29].
Histone Modification
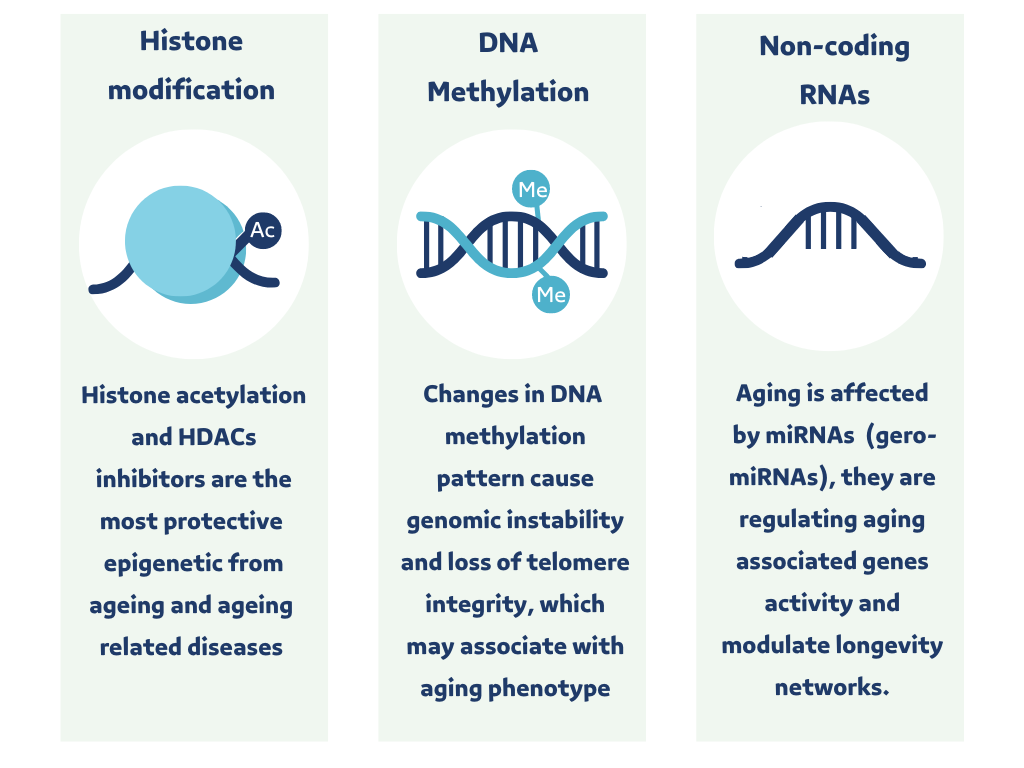
DNA in the nucleus is coiled around histone octamer and contains two copies of each histone (histone 2A (H2A), histone 2B (H2B), histone 3 (H3), and histone 4 (H4)) [27]. Histone modification leads to alteration in chromatin compaction and recruitment of transcriptional factors to modify gene expression. Acetylation of histone is the most important histone modification studied in aging research; the acetylation process is an addition of an acetyl group at the N-terminal tails of histones and makes DNA more exposed to the transcriptional factors [25, 26].
Deacetylation of histone is the removal of the acetyl group attached to the histone and compacting of the chromatin. This makes DNA less accessible to transcriptional factors and, thus, represses gene expression [30]. Histone acetylation is regulated by two types of enzymes: histone acetylases (HATs) and histone deacetylases (HDACs). HATs are enzymes for histone acetylation, and HDACs are enzymes for histone deacetylation. Histone acetylation and HDAC inhibitors are the most protective epigenetic processes from aging and aging-related diseases (Figure 1) [30].
DNA Methylation
DNA methylation (DNAm) is a process of blocking parts of DNA, which decreases transcription of the methylated sections; by adding a methyl group to the cytosine amino acid at CpG island in the DNA promoter; this modification depends on a family of enzymes known as DNA methyltransferases (DNMTs) [25, 26]. DNAm is lower in the centenarian DNA, and the methylation pattern in centenarians occurred at the gene promoter in fewer CpG islands. Changes in DNA methylation patterns could cause genomic instability and loss of telomere integrity, thus, associated with the aging phenotype (Figure 1) [10, 11].
Non-coding RNAs
Non-coding RNA (ncRNA) is an RNA molecule transcribed by DNA that does not translate into protein; ncRNA has two types short and long. The short ncRNA molecule name is microRNA (miRNA). It consists of about ~22 nucleotides and functions as a post-transcriptional regulator of gene expression by y-binding to messenger RNA, causing it to degrade or inhibit the translational process [31]. ncRNA plays a vital role in maintaining genetic stability by regulating gene expression and aging biological activity, which could provide a valuable understanding of aging and aging-related diseases [32]. (gero-miRNAs) are a miRNA class linked to aging regulating aging-associated gene activity, and modulating longevity networks at the cell and nuclear level (Figure 1) [11].
Nutri-Epigenetics and Aging
Epigenetic aging marks accumulate lifelong concerning factors; one of these factors is nutrition, which has an important environmental impact on epigenetic aging marks [16, 33]. Modification of the epigenetics related to aging has a good impact on individual health and well-being; nutrient plays an essential role in this modification, which affect transcription and translation of genes, protein production in the cells, major body organ function, and phenotype through a lifetime [16, 34-36].
Nutrient compounds include methyl donors (folic acid, vitamin B12, vitamin B6, vitamin B2, choline), which are the most important modification of the epigenetics mechanism by nutrition [34, 35]. Other essential nutrients and phytochemicals that affect epigenetics are cruciferous vegetables containing sulforaphane, soy containing genistein, grapes containing resveratrol, and tea containing epigallocatechin gallate (EGCG) [37].
Tea Catechins and Epigallocatechin Gallate
Tea (Camellia sinensis (L.) Kuntze) is the oldest and most consumed drink worldwide [38, 39]. It is a perennial shrub that belongs to Theaceae family. Moreover, Tea is an evergreen shrub native to East Asia. Classification of tea according to the fermentation process of the leaves is fermented black tea leaves, minimally fermented green tea leaves, and not fermented white tea leaves. This process makes tea types differ in their concentration and the types of phytochemicals and antioxidants in them [38-40]. Catechins (flavan-3-ol) are the most common and diversified phenolic compounds in tea and food. Catechins in the tea are epicatechin, epigallocatechin, epicatechin gallate, and EGCG is the most abundant and active catechin component in the tea, which represents about 65% of the total catechins in the tea [39].
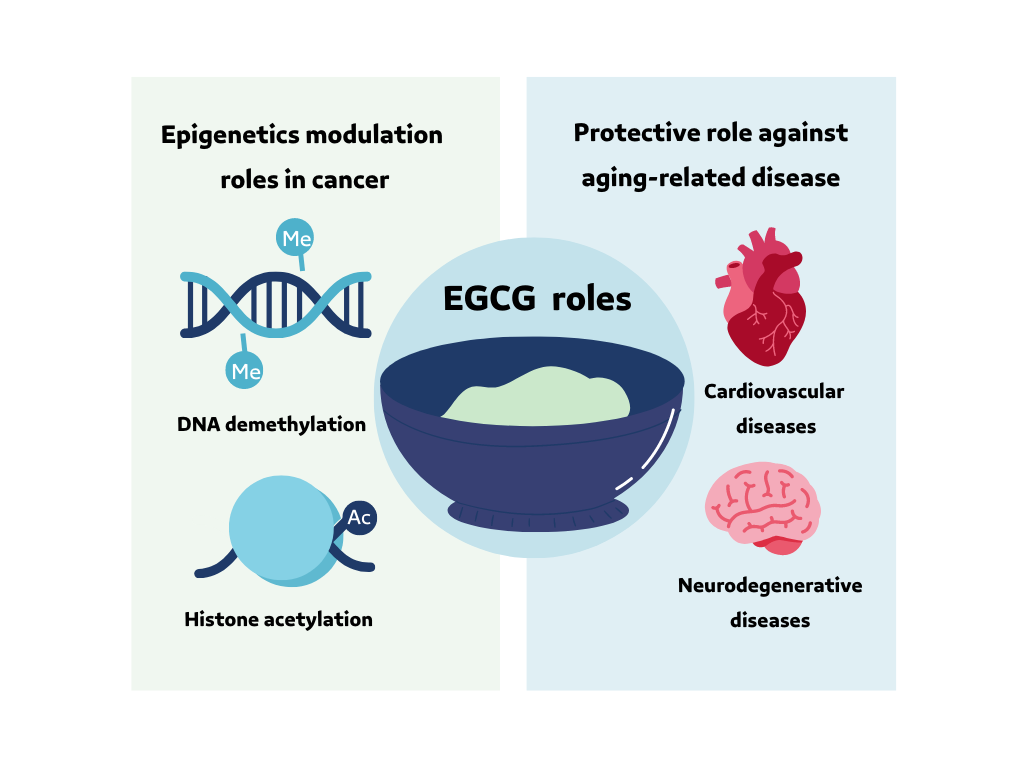
The EGCG has a beneficial role in a wide range of diseases and disorders due to it is antioxidant and anti-inflammatory activity, and it has a protective effect against aging-related diseases: neurodegenerative diseases, cardiovascular disease, and cancers (Figure 2) [41-45]. In addition, EGCG's role at the nuclear level in cancer epigenetics modulation (Figure 2) DNA demethylation and histone acetylation by inhibiting the DNMT and HDAC activities [32]. EGCG in tea has a positive and protective effect on cancer epigenetics and aging-related disease. The objective of the current review is to clarify and investigate the relationship between tea catechin EGCG and epigenetic alteration as an aging hallmark.
Materials and Methods
Study Protocol
In the current review, PRISMA Statement guidelines are followed [46]. The protocol registers in PROSPERO (CRD42021241235). Inclusion criteria were as follows: 1) Population/Animals: Mice animal model, 2) Intervention: Tea catechin compounds Epigallocatechin Gallate (EGCG), 3) Control: Younger mice and mice of the same age without intervention, 4) Study designs: Epigenetics experimental studies, full-text open-access, and 5) Outcome: Regulation of aging-related epigenetic marks on the DNA (DNA methylation, histone modification, and miRNA). Exclusion criteria were as follows: 1) Population/Animals: Human model and cell (in vitro) models, 2) Interventions: Catechins compounds other than EGCG; and catechins not extracted from the tea, 3) Study designs: Other types of studies, and 4) Outcome: Regulation of (Cancer, Alzheimer's, neurodegenerative diseases, or chronic diseases) epigenetic marks; and gene expression studies without epigenetics in the outcome excluded.
The search was conducted in the published, full-length, and open-access articles from the databases: PubMed, Science Direct, Scopus, and Google Scholar, from January 2011 to December 2020. The search terms (Table 1) and process applied in all articles parts not restricted to certain parts of the articles.
|
|
|
Figure 1. Aging epigenetics (Histone modification – DNA methylation – Non-coding RNA). The figure was produced in Canva Software (Canva Inc. Sydney, Australia) by a second author. |
|
|
|
Figure 2. EGCG beneficial roles and protective effects against aging-related disease, and nuclear role in cancer epigenetics modulation. The figure was produced in Canva Software (Canva Inc. Sydney, Australia) by a second author. |
Table 1. They used search terms and keywords in the current study.
|
Term |
The search keywords |
|
Epigenetics |
"Epigenomics, epigenetic repression, DNA methylation, histone modification, RNA, untranslated, histone code |
|
Tea catechin |
"EGCG, epigallocatechin gallate, catechin, tea, camellia sinensis, |
|
Aging |
"Aging, healthy aging, longevity" |
1 Filters were applied while searching document types (articles), open-access, and publication language (English). The process was performed and repeated for all databases.
Study Selection
Studies were collected from the databases where duplicates were deleted manually; then screening of the articles' titles, abstracts, and keywords was done for assurance. The relevant titles and abstracts were rescreened more than once. The eligibility criteria assessed in studies within the screening and full-text reading steps. Finally, studies that met our criteria were included for further investigation.
Data Extraction
Data were extracted from articles using a table containing eight characteristics according to the studies' inclusion criteria and outcome (first author, year, samples, EGCG method of intervention, duration of the intervention, tissue type, epigenetics, genes, and protein expression, antiaging effect).
Quality Assessment
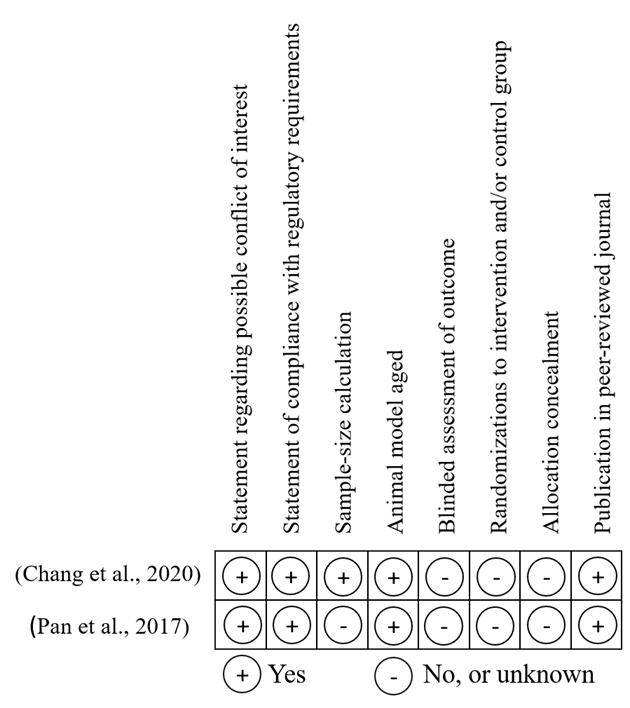
Studies quality is measured by using a modified version of the Collaborative Approach to Meta-Analysis and Review of Animal Experimental Studies (CAMARADES’) checklist depending on eight criteria (publication in the peer-reviewed journal, allocation concealment, randomizations to intervention and/or control group, blinded assessment of outcome, using of aged animal model, sample-size calculation, statement of compliance with regulatory requirements, and a statement regarding possible conflict of interest [47, 48]. The scale had a maximum score of eight and applied to all studies meeting the eligibility criteria. Studies were excluded if they scored less than four.
Results and Discussion
Study Selection
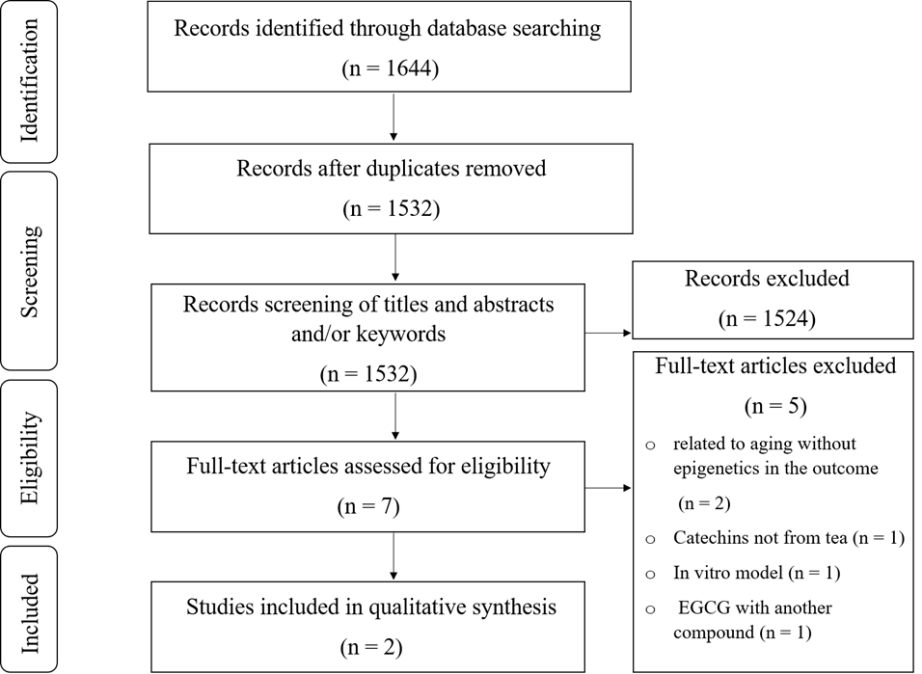
The search results in a total of 1644 articles (Figure 3), with 112 duplicate articles removed. After screening the titles, abstracts, and/or keywords, 1524 articles were excluded because they were either reviews, unrelated articles, or the intervention was by catechins other than EGCG and/or not extracted from the tea (Figure 3). After assessing the full-text articles, an additional five were excluded. Two articles met the eligibility criteria and were examined by the CAMARADES’ checklist for assessing study quality.
The overall quality of the included studies was measured, and the + indicates yes, and – indicates no or unknown if the studies met the quality measures in the checklist (Figure 4).
EGCG Modification of Aging Epigenetic
Histone Modification Through HDAC Regulation
The EGCG may increase AcH3K9 by inhibiting the expression of HDAC1 binding levels with the cTnI's proximal promoter, thus, increasing the bind levels of the transcriptional factors: GATA4 and Mef2c key cis element with cTnI gene, which will reverse the process of aging and increases the transcription of the cTnI gene, resulting in an enhancement and improvement of the cardiac function in the aged heart [49].
miRNA Expression Modification
The EGCG supplementation increases the miR-486-5p expression in skeletal muscle in aged mice compared to other mice of the same age who do not receive the EGCG supplementation. Increased miR-486-5p expression level decreases protein's expression of the phosphatase and tensin homolog (PTEN) gene [50].
Activation of serine/threonine kinase (AKT), which promote muscle growth, happens due to the decreases of PTEN protein expression, phosphorylation, and inhibition of forkhead box o1 (FoxO1) proteins which are involved in the muscle loss process, and downregulation of the transcription of muscle atrophy-related genes: muscle RING-finger protein-1 (MuRF1), and muscle atrophy F-box gene (Atrogin-1). That indicates that EGCG may contribute to a positive protein turnover in skeletal muscle and prevent age-associated muscle loss such as sarcopenia [50].
|
|
|
Figure 3. Flow diagram of search and article selection process. |
|
|
|
Figure 4. CAMARADES’ checklist [49, 50] for assessing the quality of the included studies. |
Table 2. Characteristics of the included studies. Genes and protein expression are summarized to (↓) for lower expression and (↑) for higher expression effects.
|
Antiaging effect |
Genes and protein expression |
Epigenetics |
Tissue type |
Duration of the intervention |
EGCG method of intervention |
Samples |
First author, year |
|
Positive protein turnover and prevent age-associated muscle loss. |
↓PTEN ↑AKT ↓FoxO ↓MuRF1 ↓Atrogin-1 |
↑miR-486-5p |
Skeletal muscle tissue |
8 weeks |
Chow diet containing 0.32% EGCG |
Intervention group (SAMP8/Ta Slc) 20 mice 32 weeks (8 months old) |
(Chang et al. 2020) [50] |
|
Control group (SAMR1/Ta Slc) 10 mice 32 weeks (8 months old) |
|||||||
|
Expression levels of the cTnI gene enhanced; cardiac function improved. |
↑cTnI
|
↑AcH3K9 ↓HDAC1 |
Cardiac tissue |
8 weeks |
Single dose (50 mg/kg/day) of EGCG by intraperitoneal injection |
Intervention group (SPF class c57bl/6) 16 months old mice |
(Pan et al. 2017) [49] |
|
Control group (SPF class c57bl/6) 3 months and 18 months old mice |
|||||||
|
(SAMR1/Ta Slc) senescence-accelerated mouse resistant 1, (SAMP8/Ta Slc) senescence-accelerated mouse prone 8, (AcH3K9) histone 3 lysine 9 acetylation, (HDAC1) histone deacetylase 1, (SPF) mice raised in specific pathogen-free facilities (class c57bl/6) class C57 black 6 |
|||||||
Increased life expectancy turns the world's population into aging societies; aging increases the risk of multiple disease susceptibility [2, 3]. Epigenetic alteration is an important aging hallmark [11]. Phytochemicals affect epigenetic marks, and the well-studied phytochemical compound is EGCG from tea leaves. The current review investigates the relationship between tea catechin EGCG and epigenetic alteration hallmark.
Results demonstrate a possible correlation between EGCG and muscle preservation in aging by miRNA modification; which decreases the expression of genes and proteins responsible for muscle wasting. Another role of EGCG in aging is improving cardiac diastolic function through increasing histone acetylation and HDAC lower expression (Figure 5).
In line with the quality synthesis of the article results, EGCG positively affects aging epigenetics. However, based on the findings of similar articles, it has been found that EGCG affects the epigenetics mechanism in cancer which is an aging-associated disease. EGCG has a well-studied role in histone modification by regulating HAT and HDAC enzymes. EGCG Inhibits the activity of HDAC 1 in cervical, skin, and colon cancer cells [51, 52]. Furthermore, EGCG enhances the expression of different types of miRNAs in cervical, prostate, breast, and hepatocellular carcinoma [52].
These results build on existing evidence of epigenetic alteration, EGCG reversing epigenetic mark changes due to aging through the restoration of histone acetylation through HDAC inhibitors [11]. The findings of this review suggest that EGCG stimulates histone acetylation and inhibits HDAC and this is strongly linked to the theory of epigenetic aging mark [49]. Genetic instability is a critical factor in the aging process. The miRNA is considered an essential factor in genetic stability and aging regulation. According to the aging hallmarks, there is a class of miRNAs linked to aging (gero-miRNAs) that regulate aging-associated gene activity, and according to the result, EGCG increases the miR-486-5p expression in skeletal muscles regulates aging-related genes in muscles [11, 32, 50]. These findings indicate the critical role of EGCG in epigenetic aging histone acetylation, HDAC inhibition, and miRNA expression regulation.
To our knowledge, this review is the first on the EGCG effect on epigenetic alteration as a hallmark of aging. This review focuses on epigenetic aging without another aging-related disease. The outcome is focused on the aging process according to epigenetic alteration mainly.
Key challenges and limitations of this review include that there were not enough studies for EGCG on mice covering all epigenetic modifications, such as DNA methylation, since DNA methylation is one of the important epigenetic aging marks. According to our search strategy, the open search keywords about the type of model in the systematic search were to identify all the available models, and according to this, the available models were animal models (in vivo) and cell models (in vitro) in the epigenetics studies. Open-access articles are the only type included in the search, and grey literature was not included, but we included four search databases in the literature search to minimize the effect of this obstacle on the result. There is no official quality assessment measure for epigenetic studies. Types of EGCG intervention are different. It could affect the outcome of EGCG; however, intervention through adding EGCG to diet depends on the bioavailability of EGCG for absorption in the gastrointestinal tract (GIT) is different from intervention by intraperitoneal injection allows quick and large amount reabsorption of the EGCG.
|
|
|
Figure 2. EGCG effects on epigenetic aging (Histone modification and miRNA expression). The figure was produced in Canva Software (Canva Inc. Sydney, Australia) by a second author. |
Further research in EGCG and epigenetic alteration is recommended to measure DNA methylation level concerning aging since EGCG inhibits DNA methylation by inhibiting the DNMTs in some types of oncogenes, and DNA methylation is the epigenetic clock which is strongly associated with aging [16, 51, 53] 14.58.61. Therefore, EGCG could regulate DNA methylation, inhibit DNMTs, and slow down the epigenetic clock.
An upcoming review in EGCG and epigenetic alteration should consider all types of articles other than open access only, and there is a need for quality assessment measures specific to epigenetic studies in animal and/or human model studies. Consider the type of EGCG best intervention method and its applicability in humans.
Conclusion
In summary, this review of EGCG's effects on epigenetic aging found that EGCG has a role in muscle preservation and cardiac diastolic function improvement. Muscle preservation by miRNA expression modification decreases the expression of genes and proteins responsible for muscle wasting. Cardiac diastolic function improvement is the other role of EGCG in aging through increasing histone acetylation and inhibiting HDAC 1.
There are not enough studies for EGCG on mice covering all epigenetic modifications such as DNA methylation which is one of the important epigenetic aging marks. Therefore, a future review of EGCG and epigenetic alteration should consider the need for quality assessment measures specific to epigenetic studies in animal and/or human model studies and consider the type of EGCG best intervention method and it is applicability in humans.
Acknowledgments: None
Conflict of interest: None
Financial support: None
Ethics statement: None