Pharmacophore an International Research Journal
This work investigated the therapeutic effects of flavonoid-rich extracts obtained from Hibiscus sabdariffa L. leaves in streptozotocin-induced diabetic rats. Their biochemical parameters such as redox stress biomarkers, anti-inflammatory levels, pancreatic histology, insulin concentration, lipase activity, purinergic enzyme activities, homeostasis model assessment of β-cells (HOMA-β), and homeostasis model assessment of insulin resistance (HOMA-IR) were assessed. Furthermore, the relative gene expressions of insulin receptors and GLP-1 were examined. H. sabdariffa extracts significantly (p<0.05) mitigated redox stress and inflammation in diabetic rats, as indicated by the reduction in malondialdehyde (MDA) levels, interleukin-1 beta (IL-1β), interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α), and C-reactive protein (CRP) as well as enhancement of reduced glutathione (GSH), glutathione-S-transferase (GST), catalase (CAT), and glutathione peroxidase (GPx). Pancreatic histological analysis revealed that the extracts protect against the degeneration of pancreatic acinar cells and beta cells. Moreover, they enhance pancreatic insulin concentration and reduce lipase activity in diabetic rats. In addition, the extracts positively affected purinergic enzyme activities, specifically ecto-nucleoside triphosphate diphosphohydrolase (E-NTPDase) and ATPase besides significantly improving the homeostasis model assessments of β-cells and insulin resistance. Most notably, diabetic rats treated with H. sabdariffa leaf extracts activated the relative gene expressions of insulin and GLP-1 receptors. Hence, these findings support the potential of H. sabdariffa leaf extracts as a natural therapeutic agent for managing diabetes.
Introduction
Eating a lot of prepared foods heavy in fat and sugar increases the risk of developing diabetes, dyslipidemia, and obesity. Diabetes mellitus (DM) is a significant global health problem (more than 500 million suffer from it), characterized by abnormal blood glucose levels due to insufficient or impaired insulin function, influenced by factors like physical inactivity, genetics, and lifestyle choices. It ranks among the top causes of premature death (over a million fatalities per year) [1, 2]. DM is typically divided into two main types: type-1 (T1DM) (where the body does not produce enough insulin due to the loss of insulin-producing β-cells in the pancreatic islets) and the more prevalent type 2 (T2DM) (where fat, liver, and skeletal muscle cells do not respond to insulin, reducing their ability to absorb sugar from the bloodstream) [1]. Both type 1 and type 2 diabetic models are used to study diabetes-related issues and possible treatment, even though hyperglycemia is the primary factor in both the development and progression of these adverse effects. It increases the amount of glucose autoxidation, protein glycation, and the oxidative breakdown of glycated protein that follows [2].
The synthesis and suitable secretion of insulin to regulate blood glucose levels is the pancreatic cell's primary role. Nutritional factors and hormonal regulators, such as enteroendocrine hormones (glucose-dependent insulinotropic polypeptide and glucagon-like peptide [GLP]-1), strictly control this activity [3].
Numerous cellular defense systems, including enzymatic and non-enzymatic antioxidants, control reactive oxygen species (ROS) levels. In mammalian systems, CAT, SOD, GPx, and GST are the main scavenger enzymes involved in their detoxification [4]. ROS have been shown to decrease in diabetic animals with wide variations, depending on disease duration, which could result from changes in enzyme activity over time, such as corrective increases in enzyme activity to counteract high oxidative stress or inhibitory effects. Oxidative stress, also known as elevated levels and/or insufficient elimination of ROS, can cause severe metabolic imbalance and oxidative damage to biological macromolecules. Through the process of apoptosis, which is controlled by certain functioning genes and the proteins produced by those genes, oxidative stress often causes cell death [5].
Due to the positive effects of their bioactive constituents on cellular function, as well as on insulin action, production, and resistance, some plant species have recently been appreciated in the treatment of DM. In this context, the molecular mechanisms underlying the activation of insulin and GLP-I receptors by Hibiscus sabdariffa L. (Malvaceae) leaf extracts in streptozotocin-induced Wistar rats have not been fully investigated and further data are needed to confirm their applicability in humans.
Materials and Methods
Plant material Source and Authentication
H. sabdariffa leaves were purchased from Oja-Oba market, Ado-Ekiti, Ekiti state, Nigeria. Mr. Odewo, S. A., a senior taxonomist at the Forestry Research Institute of Nigeria (FRIN), Ibadan, Nigeria, Identified and authenticated it with voucher number FHI: FHI:113742.
Chemicals, Reagents, and Enzyme Kits Used
Methanol, sulphuric acid, absolute ethanol, fructose, concentrated ammonium hydroxide, dilute ammonium hydroxide, Streptozotocin (STZ), 10 % formalin, sodium citrate buffer, and phosphate buffer were obtained from Signal Aldrich Germany, while all the reagents used were of analytical grade. Also, all the enzyme kits used were products of Randox Laboratory (Crumlin, United Kingdom).
Processing of H. Sabdariffa and Extraction of Flavonoid-Rich Extract
H. sabdariffa leaves were dried for two weeks at room temperature, then powdered with an electric blender. One gram of the sample was defatted in 80% methanol by maceration for 72 hours and then filtered through a muslin cloth. The clear filtrate obtained was concentrated using a rotary evaporator. Thereafter, 20 g of the residue was dissolved in 200 mL of 10% H2SO4 and hydrolyzed by heating in a water bath for 30 minutes at 100ºC. The mixture was placed on ice for 15 minutes for the precipitation of the flavonoid aglycones. The latter were dissolved in 50 mL of warm 95% ethanol, and filtered into a 100 mL volumetric flask which was made up to mark with 95% ethanol. This was then concentrated by a rotary evaporator. Subsequently, the filtrate was precipitated using concentrated ammonium hydroxide. The whole solution was allowed to settle and precipitate were collected and rinsed with dilute ammonium hydroxide to obtain the flavonoid extract.
Experimental Animals and Induction of Diabetes
The animals were housed in groups of five (5) and acclimatized for 7 days. Rats to be induced were given 20% fructose solution for two weeks [6]. Thereafter, 40 mg/kg body weight of streptozotocin (STZ) was administered to the animals and an equal volume of citrate buffer pH 7.4 was administered to the control group of the rat. Hence, at 72 hours of injection, animals with fasting blood glucose greater than 250 mg/dL were used in this study [7].
Experimental Design and Animal Treatment
The animals were grouped after DM induction and were placed into five groups with five rats in each group. The rats were weighed accurately and divided into different groups based on their body weight. Group I: Rats that were not induced (Normal Control); Group II: Diabetic rats without treatment (Diabetic Control); Group III: Diabetic rats administered low dose (150 mg/kg body weight) of H. sabdariffa flavonoid-rich extract leaf (LDHDFL); Group IV: Diabetic rats administered high dose (300 mg/kg body weight) of H. sabdariffaflavonoid-rich extract leaf (HDHDFL); while Group V: Diabetic rats administered 200 mg/kg of metformin (MET).
Tissue Collection and Processing
On the twenty-second day, the rats were sacrificed by cervical dislocation. Blood samples were immediately withdrawn by cardiac puncture from each rat. Their blood was collected into a plain bottle and was centrifuged for 15 minutes at 1,500 rpm. The obtained serum was kept in a refrigerator.
Also, the animal pancreas was collected and homogenized in 0.1M potassium phosphate buffer at pH 6.5. The homogenized pancreas was centrifuged at 4000 rpm for 15 minutes [8].
Determination of Malondialdehyde Level
The concentration of pancreatic MDA level was quantified according to the method of Oloyede et al. [9].
Determination of Superoxide Dismutase Activity
The method described by Misra and Fridovich [10] was used to assay for the activity of superoxide dismutase.
Determination of Glutathione-S-Transferase Activity
Glutathione transferase (GST) was measured by the method of Habig et al. [11] with modifications. For GST (CDNB) assays, the reaction medium contained 0.1M potassium phosphate buffer pH 7.5 or 6.5, 1.0 mm GSH, 1.0 mm CDNB, 1% absolute ethanol, and protein in a total volume of 1.0 mL. For GST (DCNB) assays, the reaction medium contained 0.1 m potassium phosphate buffer, pH 7.5, 5.0 or 1.0 mm GSH, 1.0 mm DCNB, 1% absolute ethanol, and protein in a total volume of 1.0 mL. The reaction, conducted at 25°C, was initiated by the addition of CDNB or DCNB, and the change in A340 or A345, respectively, was monitored for 120 s with a spectrophotometer (model DU-65, Beckman). All initial rates were corrected for the background nonenzymatic reaction. One unit of activity is defined as the formation of 1 μmol product min−1 at 25°C (extinction coefficient at 340 nm = 9.6 mm−1 cm−1 for CDNB; extinction coefficient at 345 nm = 8.5 mm−1 cm−1 for DCNB).
Determination of Catalase Activity
The method described by Beers and Sizer [12] was used to assay for the activity of catalase.
Determination of Glutathione Peroxidase Activity
Glutathione peroxidase activity was determined according to the method of Haque et al. [13].
Determination of Reduced Glutathione Level
The level of GSH in the pancreas homogenate was determined using the procedure described by Ellman [14].
Determination of Protein Carbonyl Content
Protein carbonyl content was determined according to the method of Levine et al. [15].
Determination of Interleukin-1β (IL-1β), Interleukin-6 (IL-6), Tumor Necrotic Factor-α (TNF- α), and C-Reactive Protein (CRP) Levels
These were carried out using the ELISA kit according to the method described by Bergqvist et al. [16].
Determination of Pancreatic Lipase Activity
The pancreatic lipase activity was quantified by a colorimetric assay that measures the release of p-nitrophenol as previously described [17], with minor modification. Here, p-nitrophenyl butyrate (PNPB), dissolved in acetonitrile, was employed in the enzymatic assays as PL substrate at 100 µM concentration instead of 5 mM. Aliquot (0.10 mL) of PL solution was added to the reaction mixtures. The volume was completed to 1 mL using the tris-HCl buffer before measuring the solution absorbances spectrophotometrically, at 410 nm, at a minimum of 5 time points (1 - 5 min). The reaction, maintained at 37°C, was started by adding the substrate to the reaction mixture. The release of p-nitrophenol was measured at 410 nm.
Determination of Pancreatic Insulin Concentration
Insulin levels were determined using the ELISA kit according to the method described by Thevis et al. [18].
Determination of Na+/ K+-ATPase, Ca2+/Mg2+-ATPase and Mg2+-ATPase Activities
The assays were performed according to the method described by Bewaji et al. [19].
Determination of Ecto-Nucleoside Triphosphate Diphosphohydrolase (ENTPDase) Activity
The activity was determined according to the procedure reported by Akomolafe et al. [20].
Determination of Ecto-5’-Nucleotidase Activity
The activity was determined according to the procedure reported by Freundlieb et al. [21].
Determination of HOMA-IR and HOMA-β Score
The homeostasis model assessment of insulin resistance (HOMA-IR) and beta cell function (HOMA-β) constitute a method for assessing b-cell function and insulin resistance from basal glucose and insulin concentrations according to Matthews et al. [22]. HOMA-IR and HOMA-β are also significantly associated with diabetes risk across ethnic groups. The formulas for calculating HOMA-IR and HOMA-β assessment scores are as follows:
|
HOMA-IR = [fasting insulin (µU/mL) × fasting plasma glucose (mg/dl)] / [405] |
(1) |
|
HOMA-β = [20] × [fasting insulin (µU/ml)] / [fasting plasma glucose (mg/dl) – 63] |
(2) |
Isolation of Total RNA
Total RNA was isolated from tissue samples with Quick-RNA MiniPrep™ Kit (Zymo Research). The DNA contaminant was removed following DNAse I (NEB, Cat: M0303S) treatment. The RNA was quantified at 260 nm and the purity was confirmed at 260 nm and 280 nm using A&E Spectrophotometer (A&E Lab. UK).
cDNA Conversion
One (1 μg) of DNA-free RNA was converted to cDNA by reverse transcriptase reaction with the aid of a cDNA synthesis kit based on ProtoScript II first-strand technology (New England BioLabs) in a condition of 3-step reaction: 65°C for 5 min, 42°C for 1 hr, and 80°C for 5 min [23].
PCR Amplification and Agarose Gel Electrophoresis
Polymerase chain reaction (PCR) for the amplification of the gene of interest was carried out with OneTaqR2X Master Mix (NEB) using the following primers (Inqaba Biotec, Hatfield, South Africa). PCR amplification was performed in a total of 25 μL volume reaction mixture containing cDNA, primers, and Ready Mix Taq PCR master mix. Under the following conditions: initial denaturation at 95°C for 5 min, followed by 30 cycles of amplification (denaturation at 95°C for 30 s, annealing for 30 s, and extension at 72°C for 60 s) and ending with a final extension at 72°C for 10 min. The amplicons were resolved on 1.0% agarose gel. The GAPDH gene was used to normalize the relative level of expression of each gene, and quantification of band intensity was done using “image J” software [23].
Histopathological Examination
The examination was carried out using hematoxylin and eosin (H&E) staining as described by Blume et al. [24].
Statistical Analysis
All the experimental results were shown as mean ± S.D (n = 5). Statistical significances were investigated by ANOVA followed by Tukey's multiple comparison (post-hoc test) using software (GraphPad Prism, Version 5.0). P<0.05 was fixed as statistically significant.
Results and Discussion
Effects of H. Sabdariffa Extract on Redox Stress Biomarkers in the STZ-Diabetic Rat Pancreas
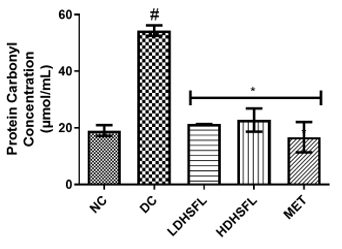
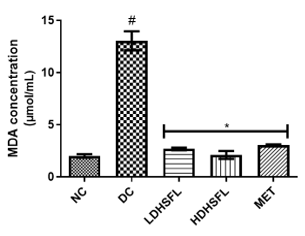
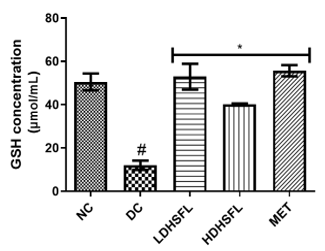
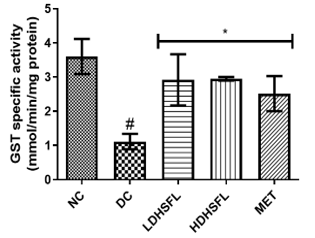
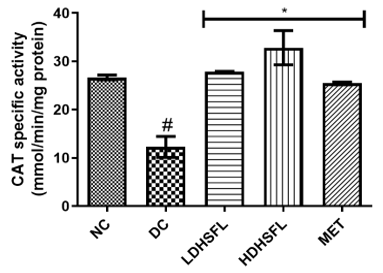
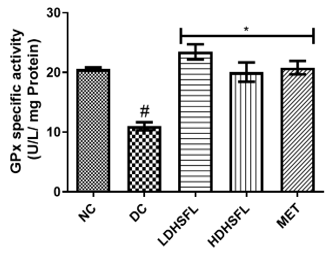
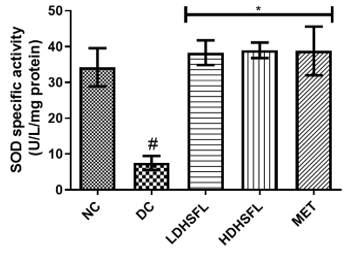
Figure 1 shows the effects of the flavonoid-rich extract obtained from H. sabdariffa leaves on the activities and/or levels of pancreatic redox stress biomarkers in STZ-diabetic rats. The data showed a significant increase (p<0.05) in both lipid peroxidation and protein degradation (measured by MDA and protein carbonyl levels, respectively) and a concomitant decrease (p<0.05) in the activities of antioxidant enzymes (glutathione-S-transferase, GST; catalase, CAT; glutathione peroxidase, GPx; and superoxide dismutase, SOD) and in the level of reduced glutathione (GSH) in the untreated diabetic group (DC) compared to the normal group. However, these misdemeanors were significantly (p<0.05) attenuated in a dose-dependent manner by the administration of both low and high doses of the extract to closely resemble the group treated with standard metformin, highlighting its potent antioxidant properties, capable of mitigating oxidative stress and excess free radicals associated with diabetes, in line with previous research in this field [25].
In the study, normal control rats without diabetes did not experience an increase in antioxidant enzyme activities, as they did not produce ROS. In contrast, positive control diabetic rats exposed to STZ faced a different situation due to damaged pancreatic β-cells, resulting in insulin deficiency and elevated ROS levels [26]. Increasing the activity and gene expression of antioxidant enzymes is critical to managing such stressful conditions. Similar to this research, STZ-induced diabetic rats have previously shown increased MDA activity in the pancreas, which can harm various components, including enzymes, phospholipids, proteins, nucleic acids, and cell membranes [27]. Patients with diabetes experience significant oxidative stress due to increased oxidation products and reduced free radical removal. Protection of the organism from free radical damage involves reducing lipid peroxidation products, preservation of cell membrane integrity, and maintenance of normal cellular functions. The integrity of the cell membrane structure is particularly crucial for the proper functioning of insulin receptors, as they are a type of cell membrane receptor [28].
|
|
|
|
|
a) |
b) |
c) |
|
|
|
|
|
d) |
e) |
f) |
|
|
||
|
g) |
||
|
Figure 1. Redox stress biomarkers in STZ-diabetic rats after treatment with flavonoid-rich extract from H. sabdariffa leaves. Each value is the mean of eight determinations ± SD. # p<0.05 vs NC, * p<0.05 vs DC. NC, Normal Control; DC, Diabetic Control; LDHSFL, STZ-diabetic rats treated with a low dose (150 mg/kg body weight) of extract; HDHSFL, STZ-diabetic rats treated with a high dose (300 mg/kg body weight) of extract; MET, STZ-diabetic rats treated with 200 mg/kg of metformin; MDA; malondialdehyde; GSH, reduced glutathione; GST, glutathione-S-transferase; CAT, catalase; GPx, glutathione peroxidase; SOD, superoxide dismutase. |
||
Effects of H. Sabdariffa Extract on Pro-Inflammatory Biomarkers in the STZ-Diabetic Rat Pancreas
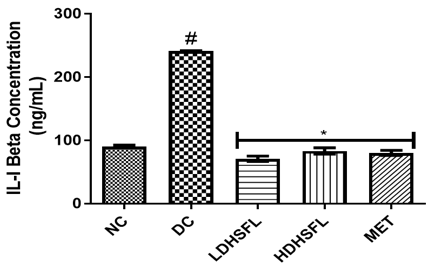
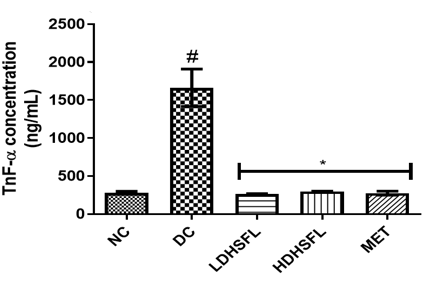
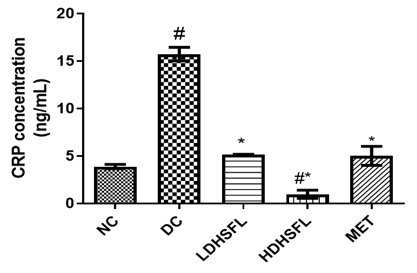
As shown in Figure 2, the administration of STZ steered to a substantial (p<0.05) increase in the levels and concentration of pancreas pro-inflammatory markers such as interleukin-1β (IL-1β), interleukin-6 (IL-6), tumor necrotic factor-α (TnF-α) and C-reactive Protein (CRP) in the diabetic untreated (DC) rats as compared with the normal control rats (NC), in line with those of previous studies [29]. This links to the well-documented concept that pro-inflammatory cytokines, particularly TNF-α, can induce insulin resistance. In cases of obesity, where adiponectin levels decrease as fat levels rise, various cytokines, including TNF-α, IL-6, and others, are secreted by adipocytes and activated macrophages, contributing to a sub-acute inflammatory state associated with obesity, insulin resistance, and the development of type-2 diabetes [30]. Understanding these inflammation-induced pathways is crucial for addressing the complex relationship between obesity, inflammation, and insulin resistance. STZ-induced inflammation can also contribute to pancreas-inflammation, a common complication in diabetes. C-reactive protein (CRP) serves as an essential inflammation marker in the body, with a growing association with T2DM due to low-grade inflammation contributing to insulin resistance. Elevated CRP levels are more commonly linked to increased cardiovascular disease risk, particularly in T2DM, leading to its monitoring in high-risk diabetic patients [31].
The deleterious increases in pro-inflammatory biomarkers were significantly (p<0.05) counteracted by post-administration of the flavonoid-rich extract of H. sabdariffa (150 mg/kg body weight and 300 mg/kg body weight) and 200 mg/kg body weight of metformin as standard drug. The administration of the extract inhibited the release of TNF-α, IL-1β, IL-6, and CRP, probably influencing insulin secretion directly or indirectly through the regulation of hepatic enzymes responsible for glucose metabolism, similar to metformin treatment, indicating its potential anti-inflammatory capacity and its role in reducing diabetes-related pancreas-inflammation [32].
Effects of H. Sabdariffa Extract on Histoarchitectural Modifications in the STZ-Diabetic Rat Pancreas (Hippocampus)
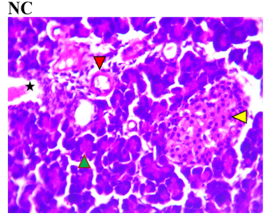
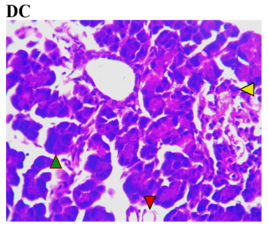
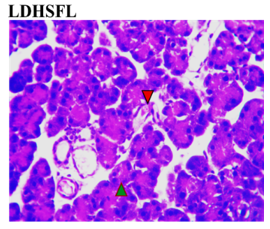
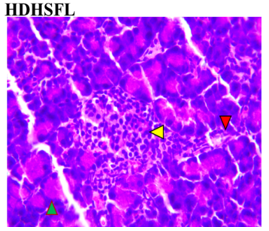
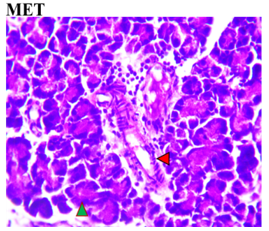
Figure 3 shows the effects of the flavonoid-rich extract from H. sabdariffa on the pancreas of STZ-induced diabetic rats. As expected, distinct deleterious histological changes characteristic of liver injury, such as severe loss of acidophilic pancreatic acinar cells and substantial degeneration of insulin-producing β-cells in the islets of Langerhans, leading to decreased insulin production and hyperglycemia, were observed in the photomicrograph of the untreated diabetic group (DC) compared to the normal group (NC).
As previously documented, this situation is associated with other histopathological changes, crucial for understanding the pathophysiology of diabetes and its related complications, to underline the structural and cellular transformations resulting from chronic hyperglycemia and STZ toxicity [33]. Signs of inflammation and fibrosis are prevalent, indicative of an immune response to STZ-induced damage, and vascular sclerosis may affect blood flow and nutrient supply in the pancreas. Amyloid deposits may occur in islets, and ductal changes may impact exocrine pancreatic function [34].
In the study, however, after careful microphotography examination, the initially observed disorders were significantly restored to near-normal conditions by the administration of both low and high doses of the H. sabdariffa extract as well as the standard compound. Treatments with H. sabdariffa extract reversed the effect on pancreatic acinar cells and β-cells, suggesting a partial remission mechanism [25].
|
|
|
|
a) |
b) |
|
|
|
|
c) |
d) |
|
Figure 2. Pro-inflammatory markers in STZ-diabetic rats after treatment with flavonoid-rich extract from H. sabdariffa leaves. Each value is the mean of eight determinations ± SD. # p<0.05 vs NC, * p<0.05 vs DC. NC, Normal Control; DC, Diabetic Control; LDHSFL, STZ-diabetic rats treated with a low dose (150 mg/kg body weight) of extract; HDHSFL, STZ-diabetic rats treated with a high dose (300 mg/kg body weight) of extract; MET, STZ-diabetic rats treated with 200 mg/kg of metformin; IL-1 beta: Interleukin-1 beta; IL-6, Interleukin-6; TnF-α, tumor necrosis factor- α; CRP, C-reactive protein. |
|
|
|
|
|
|
|
a) |
b) |
c) |
|
|
|
|
||
|
d) |
e) |
||
|
Figure 3. Pancreatic section histoarchitecture in STZ-diabetic rats after treatment with flavonoid-rich extract from H. sabdariffa leaves. H&E stain at x800 magnification. NC, Normal Control; DC, Diabetic Control; LDHSFL, STZ-diabetic rats treated with a low dose (150 mg/kg body weight) of extract; HDHSFL, STZ-diabetic rats treated with a high dose (300 mg/kg body weight) of extract; MET, STZ-diabetic rats treated with 200 mg/kg of metformin. NC shows a normal acidophilic pancreatic acinar cell (green arrowhead), normal blood vessels (star), and evenly distribution of β-cells in the endocrine region; DC shows severe loss of acidophilic pancreatic acinar cell (green arrowhead) and severe degeneration of β-cells in the endocrine region; LDHSFL shows moderate degeneration of exocrine pancreatic acinar cell (green arrowhead) and moderate degeneration of endocrine region; HDHSFL shows a normal acidophilic pancreatic acinar cell (green arrowhead) and evenly distribution of β-cells in the endocrine region; MET shows moderate degeneration of exocrine pancreatic acinar cell (green arrowhead) and degeneration of the endocrine region. |
|||
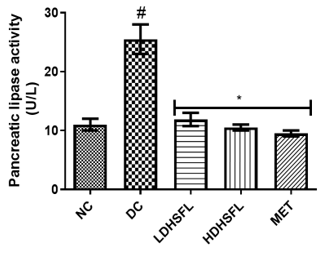
Effects of H. Sabdariffa Extract on Insulin and Lipase in the STZ-Diabetic Rat Pancreas
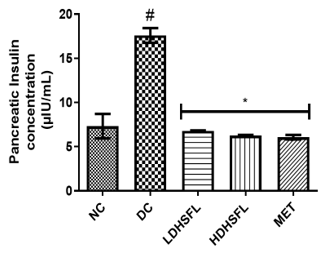
Administration of STZ to untreated diabetic rats resulted in a significant increase (p<0.05) in pancreatic lipase activity with a concomitant increase in pancreatic insulin concentrations above normal, indicating increased lipolysis and dysfunction of insulin signal [35].
As shown in Figure 4, treatment with H. sabdariffa extracts (low and high doses) was able to significantly reverse (p<0.05), in a dose-dependent manner, and finally normalize this negative trend, similarly to metformin. These results suggest the extract's potential for pancreas protection and improving insulin sensitivity, confirming previously published data [36].
|
|
|
|
a) |
b) |
|
Figure 4. Pancreatic insulin concentration and lipase activity in STZ-diabetic rats after treatment with flavonoid-rich extract from H. sabdariffa leaves. Each value is the mean of eight determinations ± SD. # p<0.05 vs NC, * p<0.05 vs DC. NC, Normal Control; DC, Diabetic Control; LDHSFL, STZ-diabetic rats treated with a low dose (150 mg/kg body weight) of extract; HDHSFL, STZ-diabetic rats treated with a high dose (300 mg/kg body weight) of extract; MET, STZ-diabetic rats treated with 200 mg/kg of metformin. |
|
Understanding the role of pancreatic insulin levels and lipase activity in diabetes is crucial for effective management. In T1DM, the immune system destroys insulin-producing cells, causing severe insulin deficiency, while T2DM involves initial insulin production but resistance in body cells causes high blood sugar levels and related problems [37]. T1DM is managed with insulin replacement through injections or pumps, while T2DM management focuses on improving insulin sensitivity, blood glucose regulation, and, if necessary, insulin-stimulating medications, all complemented by lifestyle changes such as a healthy diet and exercise [38]. Lipase, essential for the digestion of fats, may be impaired in diabetes due to digestive complications related to the condition. Dietary adjustments tailored to diabetes type can help manage lipase function, and medications may be prescribed to alleviate digestive symptoms [39].
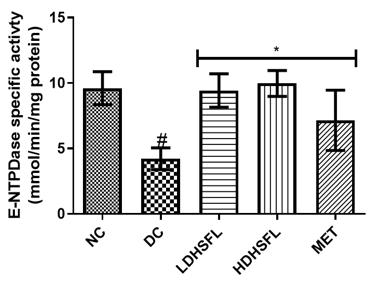
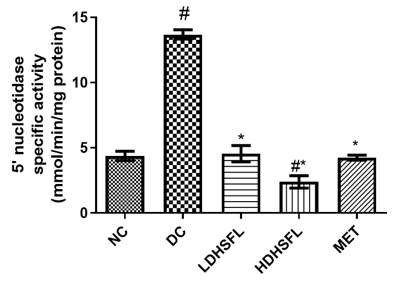
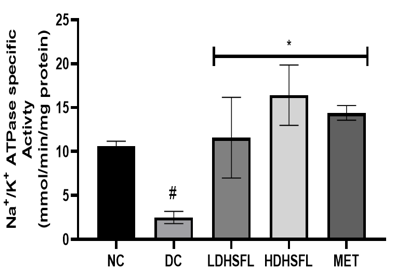
Effects of H. Sabdariffa Extract on Purinergic Enzyme Activities in the STZ-Diabetic Rat Pancreas
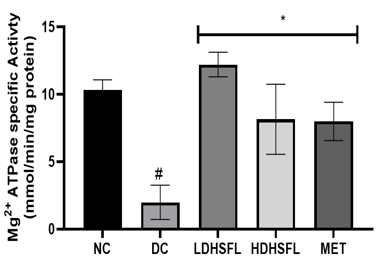
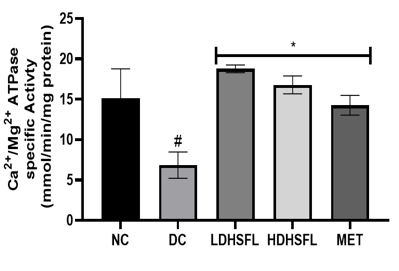
As shown in Figure 5, STZ administration also led to a significant decrease (p<0.05) in the activities of purinergic enzymes such as ecto-nucleoside triphosphate diphosphohydrolase, E-NTPDase, Na+/K+ATPase, Ca2+/Mg2+ATPase and Mg2+ATPase, along with a significant increase (p<0.05) in 5'-nucleotidase activity, in untreated diabetic rats compared to those in the normal group. These enzymes typically play a role in regulating purinergic signaling and energy production [40]. E-NTPDases break down ATP and ADP into AMP, influencing processes such as neurotransmission and immune responses, while ATPase converts ATP into ADP and Pi releasing energy for metabolic activities. The combined effect of these enzymatic activities results in a significant decrease in nucleotide levels and energy in the pancreas [41].
Importantly, administration of H. sabdariffa (150 mg/kg body weight and 300 mg/kg body weight) effectively mitigated (p<0.05) purinergic enzyme abnormalities, similar to the effects of metformin (200 mg /kg body weight). The extract significantly increased (p<0.05) E-NTPDase and ATPase activity. These results suggest that it has the potential to improve purinergic signaling in diabetic rats and may influence their neurological function [42].
|
|
|
|
|
|
a) |
b) |
c) |
|
|
|
|
||
|
d) |
e) |
||
|
Figure 5. Purinergic enzyme activities in STZ-diabetic rats after treatment with flavonoid-rich extract from H. sabdariffa leaves. Each value is the mean of eight determinations ± SD. # p<0.05 vs NC, * p<0.05 vs DC. NC, Normal Control; DC, Diabetic Control; LDHSFL, STZ-diabetic rats treated with a low dose (150 mg/kg body weight) of extract; HDHSFL, STZ-diabetic rats treated with a high dose (300 mg/kg body weight) of extract; MET, STZ-diabetic rats treated with 200 mg/kg of metformin; E-NTPDase, ecto-nucleoside triphosphate diphosphohydrolase. |
|||
Effect of H. Sabdariffa Extract on Homeostasis Model Assessment of β-Cells and Insulin Resistance in the STZ-Diabetic Rat Pancreas
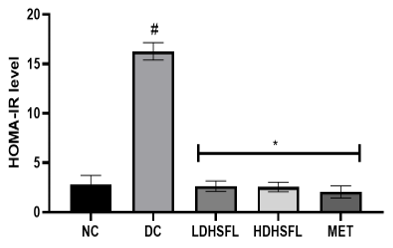
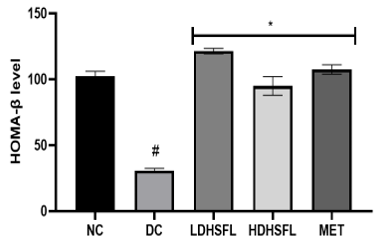
This study found that induction of diabetes in rats using STZ results in a significant increase (p<0.05) in the level of homeostasis model assessment of insulin resistance (HOMA-IR), an equally significant reduction (p<0.05) in the level of homeostasis model assessment of β-cell (HOMA-β) and higher levels of pancreatic insulin in untreated diabetic rats compared to those in the normal group (Figure 6). The disastrous inconsistencies in HOMA-IR and HOMA-B scores were significantly improved (p<0.05) after administration of H. sabdariffa extract (150 mg/kg body weight and 300 mg/kg body weight), similar to what happened with the standard drug metformin (200 mg/kg body weight) (Figure 6). Rats treated with the extract after STZ administration showed a decrease in HOMA-IR (Figure 6). These data confirm the results of previous research suggesting that the plant extracts can improve β-cell recovery and insulin resistance [43, 44].
|
|
|
|
a) |
b) |
|
Figure 6. Homeostasis Model Assessment of β-cells (HOMA-β) and Homeostasis Model Assessment of Insulin Resistance (HOMA-IR) in STZ-diabetic rats after treatment with flavonoid-rich extract from H. sabdariffa leaves. Each value is the mean of eight determinations ± SD. # p<0.05 vs NC, * p<0.05 vs DC. NC, Normal Control; DC, Diabetic Control; LDHSFL, STZ-diabetic rats treated with a low dose (150 mg/kg body weight) of extract; HDHSFL, STZ-diabetic rats treated with a high dose (300 mg/kg body weight) of extract; MET, STZ-diabetic rats treated with 200 mg/kg of metformin. |
|
Effect of H. Sabdariffa Extract on Relative Gene Expressions of IR and GLP-1 in the STZ-Diabetic Rat Pancreas
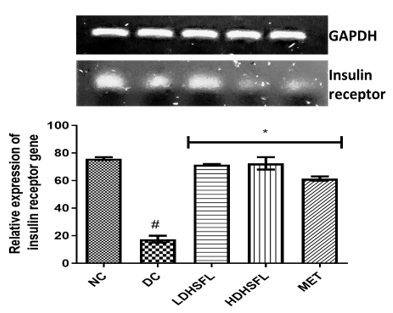
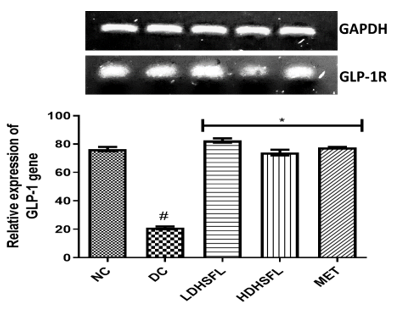
Finally, Figure 7 reveals the gene expression patterns of insulin receptor (IR) and glucagon-like peptide (GLP-1) genes in the pancreas of STZ-induced diabetic rats and the resulting restorative effect of the flavonoid-rich extract of H. sabdariffa. Using glycemic acid-3-phosphate dehydrogenated hydrochloride (GAPDH) as a housekeeping gene, a significant downregulation (p<0.05) of the two genes was initially observed in the DC group compared to the NC group. In line with previous literature [36, 45], the administration of the extract significantly upregulated (p<0.05) and restored their gene expression in a dose-dependent manner, comparable to standard metformin, by hypothetically binding to the promoter/enhancer region of IL and GLP-1 and inhibiting their repressor genes.
|
|
|
|
a) |
b) |
|
Figure 7. Relative gene expressions of insulin receptor and GLP-1 levels in STZ-diabetic rats after treatment with flavonoid-rich extract from H. sabdariffa leaves. Each value is the mean of eight determinations ± SD. # p<0.05 vs NC, * p<0.05 vs DC. NC, Normal Control; DC, Diabetic Control; LDHSFL, STZ-diabetic rats treated with a low dose (150 mg/kg body weight) of extract; HDHSFL, STZ-diabetic rats treated with a high dose (300 mg/kg body weight) of extract; MET, STZ-diabetic rats treated with 200 mg/kg of metformin; GLP-1, glucagon-like peptide. Note that the grouping of gels/blots was cropped from different gels. |
|
Gene regulation plays a critical role in diabetes management by influencing the up-and down-regulation of genes that control insulin production, glucose metabolism, and other key processes. Modifying gene expression through lifestyle changes, drugs, and emerging therapies offers the potential to improve blood sugar management, increase insulin sensitivity, and reduce the risk of diabetes-related complications [46]. In the context of STZ-induced diabetes in rats, drug administration leads to dramatic alterations in gene expression, primarily affecting IR and GLP-1. The reduction in IR is primarily associated with the toxic impact of STZ on insulin-producing pancreatic β-cells. The damage impairs insulin production and subsequently reduces gene expression. STZ, which is responsible for the overall disruption of pancreatic function, also affects the expression of the GLP-1 gene, an important regulator of blood glucose levels [47]. These gene expression changes reflect STZ-induced pancreatic damage and insulin deficiency, highlighting the importance of managing these changes in diabetes treatment to restore normal insulin and GLP-1 function and thus improve blood glucose regulation. The major promoters of liver glycogen synthesis and inhibitors of its secretion are key insulin receptor substrate molecules, typically expressed in hepatocytes and pancreatic cells. The obtained data suggest that flavonoid-rich extract of H. sabdariffa leaves can improve insulin signaling in diabetic rats by modulating key insulin genes [48].
Conclusion
In summary, this study underscores the remarkable therapeutic potential of H. sabdariffa leaf extracts in addressing streptozotocin-induced diabetes in Wistar rats. The research reveals a broad spectrum of effects, encompassing both molecular and physiological dimensions of diabetes management. The samples demonstrate a significant capacity to alleviate oxidative stress and inflammation. Moreover, they play a crucial role in preserving pancreatic integrity and enhancing the distribution of b-cells, offering promise for improved diabetes management. These extracts positively influence insulin concentration, lipid metabolism, purinergic enzyme activities, and homeostasis model assessments of β-cells and insulin resistance. Most notably, they activate gene expressions of IR and GLP-1, shedding light on their potential to enhance glycemic control. Overall, the findings provide a robust foundation for further research and clinical exploration of H. sabdariffa as a multifaceted approach to managing diabetes and its associated complications.
Acknowledgments: None
Conflict of interest: None
Financial support: None
Ethics statement: This study was carried out on animals and all experimental methods were approved by FUOYE Faculty of Science Ethics Committee with ethics number FUOYEFSC 201122-REC2022/008.